TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
公開番号
2025065059
公報種別
公開特許公報(A)
公開日
2025-04-17
出願番号
2024173430
出願日
2024-10-02
発明の名称
細胞質送達ペプチド
出願人
国立大学法人京都大学
代理人
個人
,
個人
主分類
C07K
14/00 20060101AFI20250410BHJP(有機化学)
要約
【課題】本開示の1つの目的は、細胞質送達ペプチドを提供することである。
【解決手段】N末端から順に、酸性アミノ酸ドメイン、マストパランドメインおよび塩基性アミノ酸ドメインを含むか、または、N末端から順に、塩基性アミノ酸ドメイン、マストパランドメインおよび酸性アミノ酸ドメインを含み、酸性アミノ酸ドメインが1個~3個の酸性アミノ酸からなり、マストパランドメインが特定のアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなり、塩基性アミノ酸ドメインが12個以上の塩基性アミノ酸からなる、細胞質送達ペプチドを提供する。
【選択図】なし
特許請求の範囲
【請求項1】
細胞質送達ペプチドであって、
N末端から順に、酸性アミノ酸ドメイン、マストパランドメインおよび塩基性アミノ酸ドメインを含むか、または、N末端から順に、塩基性アミノ酸ドメイン、マストパランドメインおよび酸性アミノ酸ドメインを含み、
酸性アミノ酸ドメインが1個~3個の酸性アミノ酸からなり、
マストパランドメインが配列番号1のアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなり、
塩基性アミノ酸ドメインが12個以上の塩基性アミノ酸からなる、
細胞質送達ペプチド。
続きを表示(約 610 文字)
【請求項2】
N末端から順に、酸性アミノ酸ドメイン、マストパランドメインおよび塩基性アミノ酸ドメインを含む、請求項1に記載のペプチド。
【請求項3】
塩基性アミノ酸ドメインが、ヒスチジン、リシンおよびアルギニンから選択される12個以上の塩基性アミノ酸からなる、請求項1に記載のペプチド。
【請求項4】
マストパランドメインが配列番号1のアミノ酸配列を含む、請求項1に記載のペプチド。
【請求項5】
酸性アミノ酸ドメインが少なくとも1個のD-アミノ酸を含む、請求項1に記載のペプチド。
【請求項6】
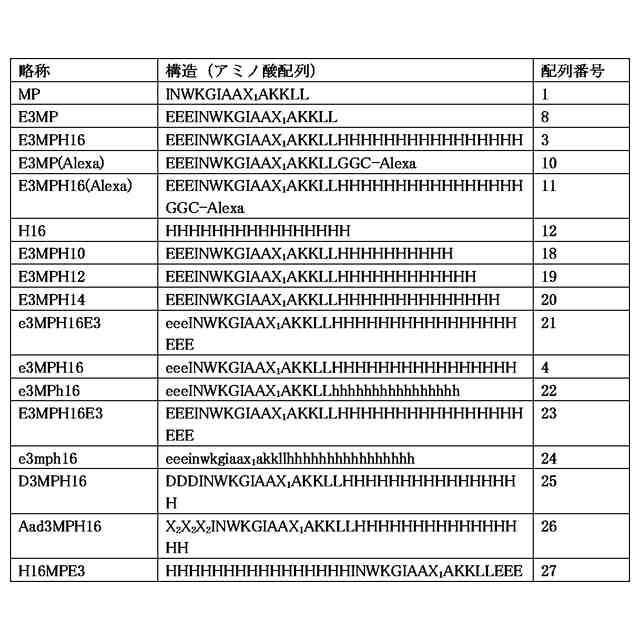
配列番号3、4および19~27のいずれかのアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなる、請求項1に記載のペプチド。
【請求項7】
配列番号3、4および19~27のいずれかのアミノ酸配列からなる、請求項1に記載のペプチド。
【請求項8】
配列番号3または4のアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなる、請求項1に記載のペプチド。
【請求項9】
配列番号3または4のアミノ酸配列からなる、請求項1に記載のペプチド。
【請求項10】
システインを含むSS結合配列がC末端に結合している、請求項1に記載のペプチド。
(【請求項11】以降は省略されています)
発明の詳細な説明
【技術分野】
【0001】
本開示は、細胞質送達ペプチドに関するものである。
続きを表示(約 6,200 文字)
【背景技術】
【0002】
抗体などの高分子の医薬品候補物質が増加しているが、細胞膜を透過できないものが多く、その標的は細胞外分子や膜タンパク質の細胞外領域に限定される。細胞内にある分子を標的にするには、これらの物質を目的の細胞内に送り込む技術が必要である。既存の細胞質送達ペプチドは、例えば、特許文献1、非特許文献1および2などに開示されている。
【先行技術文献】
【特許文献】
【0003】
特許第6202707号公報
【非特許文献】
【0004】
Azuma, Y. et al. (2018) Angew. Chem. Int. Ed. 57, 12771-12774
Iwasaki, T. et al. (2015) J. Control. Release 210, 115-124
【発明の概要】
【発明が解決しようとする課題】
【0005】
本開示の目的は、改良された細胞質送達ペプチドを提供することである。
【課題を解決するための手段】
【0006】
ある態様では、本開示は、細胞質送達ペプチドであって、
N末端から順に、酸性アミノ酸ドメイン、マストパランドメインおよび塩基性アミノ酸ドメインを含むか、または、N末端から順に、塩基性アミノ酸ドメイン、マストパランドメインおよび酸性アミノ酸ドメインを含み、
酸性アミノ酸ドメインが1個~3個の酸性アミノ酸からなり、
マストパランドメインが配列番号1のアミノ酸配列と90%以上の同一性を有するアミノ酸配列からなり、
塩基性アミノ酸ドメインが12個以上の塩基性アミノ酸からなる、
細胞質送達ペプチドを提供する。
【0007】
ある態様では、本開示は、タンパク質に細胞質送達ペプチドを結合させる方法であって、
(1)システインを含むSS結合配列がC末端に結合している上記ペプチドを提供すること、
(2)(1)のペプチドとG
n
-(リンカー)-C(nは2以上の整数である)の構造を含むペプチドを接触させ、ジスルフィド結合を形成させること、
(3)LPXTG(Xは任意のアミノ酸である)を含むアミノ酸配列がC末端に結合しているタンパク質を提供すること、
(4)(2)で得られたペプチドと(3)のタンパク質をソルターゼにより反応させること、
を含む方法を提供する。
【0008】
ある態様では、本開示は、上記ペプチドをコードするポリヌクレオチドを提供する。
ある態様では、本開示は、上記ポリヌクレオチドを含むベクターを提供する。
ある態様では、本開示は、上記ポリヌクレオチドまたは上記ベクターを含む細胞を提供する。
ある態様では、本開示は、細胞内に物質を送達するための、上記ペプチドを含む組成物を提供する。
ある態様では、本開示は、細胞内に物質を送達する方法であって、上記ペプチドおよび細胞内に送達すべき物質を細胞にインビトロで接触させることを含む、方法を提供する。
【発明の効果】
【0009】
本開示により、改良された細胞質送達ペプチドが提供される。
【図面の簡単な説明】
【0010】
本開示の背景技術。MP(配列番号1)からE3MP(配列番号8)に構成を変更することにより、毒性が低下するものの、細胞内への取り込み効率も低下してしまう。さらに、E3MPからE3-Dpa-MP(配列番号9)に構成を変更することにより、毒性は上がるものの、細胞内への取り込み効率は向上する。
本開示の細胞質送達ペプチドのデザイン。本開示の細胞質送達ペプチドによれば、低毒性と高い細胞内への取り込み効率が実現される。
E3MPとE3MPH16のHT1080細胞における細胞毒性。MP(空白のダイヤモンド;◇)、E3MP(空白の三角形;△)、およびE3MPH16(塗りつぶしの円形;●)を記載の濃度で18時間HT1080細胞に処理した際の細胞毒性。E3MPH16は100μMでも細胞毒性を示さなかった。
E3MPとE3MPH16のHT1080細胞における細胞内取り込み。10μMのE3MPGGC(Alexa488)(E3MP(Alexa))とE3MPH16GGC(Alexa488)(E3MPH16(Alexa))を1時間HT1080細胞に処理した際の細胞内取り込みのフローサイトメトリーによる定量。E3MPH16はE3MPと比べて40倍の細胞内への取り込み促進が可能になった。
E3MPH16によるDex-488の細胞質送達とエンドソーム脱出促進。(左)200μg/mL Dex-488と10μMの記載のペプチドで18時間処理したHT1080細胞の共焦点顕微鏡画像。(右)左の画像において細胞質でのDex-488シグナルを有する細胞の割合。E3MPH16は血清存在下であっても蛍光標識されたデキストランを細胞質に送達した。
10μM E3MPH16を200μg/mLのDex-488とともに18時間様々な細胞に処理した際の共焦点顕微鏡画像。E3MPH16は様々な細胞種で細胞質にデキストランを送達した。
E3MPH16によるNLS-GFPの細胞質送達。5μMのNLS-GFPと10μMのE3MPH16を添加して、18時間インキュベートしたHT1080細胞の共焦点顕微鏡画像。E3MPH16によって、核内移行シグナルを付与したGFPを核内に導入することが可能になった。
E3MPH16による機能性タンパク質の細胞質送達。(上)Cre-loxP組換えアッセイの模式図。(下)5μMCreまたは5μMCreおよび10μME3MPもしくはE3MPH16を24時間処理したloxP-レポーターHEK293細胞における遺伝子組換え効率。E3MPH16によって細胞内に送達された組換えタンパク質Creは核内に移行して染色体上のloxpサイトを認識して染色体DNAの組み換えを高効率で引き起こした。
E3MPH16による機能性タンパク質の細胞質送達。500μg/mLの抗GFP抗体とE3MP16を18時間処理し、固定後に免疫染色したHT1080細胞の共焦点顕微鏡画像。E3MPH16によって細胞内に送達された抗GFP抗体は、細胞内で分解や変性などされることなく、細胞内のGFPに特異的に結合した。
E3MPH16による機能性タンパク質の細胞質送達。500μg/mLまたは167μg/mLの抗Ras抗体と10μME3MPH16を24時間処理したHT1080細胞における細胞増殖抑制効果。E3MPH16によって細胞内に導入された抗Ras抗体は細胞内のRasタンパク質に結合して細胞増殖を抑制した。
ソルターゼによるタンパク質とペプチドのコンジュゲート作製法の模式図。
ソルターゼによるタンパク質とペプチドのコンジュゲート作製。(左上)非還元SDS-PAGEによるタンパク質とペプチドのコンジュゲートの確認。(下)4μMのNLS-GFPとE3MPH16のコンジュゲートをHT1080細胞に添加して18時間インキュベート後の共焦点顕微鏡画像。NLS-GFPとE3MPH16のコンジュゲートにより、NLS-GFPは高効率で核内に送達された。
(左)10μMの指定されたペプチドを200μg/mLのDex-488とともに18時間処理したHT1080細胞の共焦点顕微鏡画像。(右)左の画像に基づき細胞質内に拡散したDex-488シグナルを持つ細胞の割合。E3MPのC末側に付加するポリヒスチジンのヒスチジン残基の数は、12個になると細胞内への送達効率が上昇した。
E3MPH16のN末をD体にすることで活性が改善向上。10μMの指定されたペプチドを200μg/mLのDex-488とともに18時間処理したHT1080細胞の共焦点顕微鏡画像。E3MPH16のN末側のグルタミン酸(E)残基をD体にすることにより(図中でD体のアミノ酸残基を小文字で表記する。)、細胞内への送達効率が向上した。
E3MPH16のN末をD体にすることで活性が改善向上。5μMCreまたは5μMCreおよび10μMの指定されたペプチドを24時間処理したloxP-レポーターHEK293細胞における遺伝子組換え効率。E3MPH16のN末側のグルタミン酸(E)残基をD体にすることにより(図中でD体のアミノ酸残基を小文字で表記する。)、組換えタンパク質Creの細胞内への送達とこれに続く核内移行が促進され、染色体上のloxpサイト認識を介した染色体DNAの組み換えを高効率で引き起こした。
担癌マウスにおける局所投与によるNLS-GFPのE3MPH16を介した細胞質送達(上)大腸-26ゼノグラフト腫瘍モデルマウスへの局所投与によるE3MPH16によるNLS-GFPの配達の模式図。(下)大腸-26ゼノグラフト腫瘍モデルマウスに5μMのNLS-GFP、5μMのNLS-GFPおよびE3MPH16、または4μMのNLS-GFP-E3MPH16コンジュゲートを局所投与してから24時間後、組織を収集し、パラフィンスライドを準備した。核をDAPIで染色し、それぞれの蛍光は共焦点顕微鏡によって観察した。画像中の白いボックスは拡大表示された領域を示す。NLS-GFPによりE3MPH16はがん細胞の核内に移行した。
E3MPH16によるDex-488の細胞質送達とエンドソーム脱出促進。10μMのE3MP16と200μg/mLのDex-488で記載の時間処理したHT1080細胞の共焦点顕微鏡画像。E3MPH16はエンドソーム経路から被送達物質を細胞内に送達していることが判明した。
200μg/mLのDex-488および10μMのE3MPH16で処理され、1時間のインキュベート後に洗浄されたHT1080細胞の共焦点顕微鏡画像(右パネル)およびその後さらに2時間インキュベートしたHT1080細胞の共焦点顕微鏡画像(左パネル)。E3MPH16はエンドソームから被送達物質を細胞内に送達していることが判明した。
E3MPH16の細胞質送達に関わる細胞内取り込み経路。クラスリン依存性エンドサイトーシス、カベオラ依存性エンドサイトーシスやマクロピノサイトーシスに対するエンドサイトーシス阻害剤で処理したHT1080細胞において10μMのE3MPH16(Alexa)を添加し、1時間インキュベートした際の細胞内取り込み量。E3MPH16による被送達物質の細胞内送達にはエンドサイトーシスが関与することが判明した。
E3MPH16の細胞質送達に関わる細胞内取り込み経路。(左)エンドサイトーシス阻害剤を30分、HT1080細胞に処理したのちに、エンドサイトーシス阻害剤と10μME3MPH16と200μg/mLDex-488を共にHT1080細胞に添加し、1時間インキュベートした際の共焦点顕微鏡画像。(右)左の画像において細胞質全体にDex-488シグナルが拡散した細胞の割合。E3MPH16によるエンドサイトーシスを介した被送達物質の細胞内送達には、マクロピノおよびカベオラ経路が関与することが判明した。
E3MPH16によるDex-488の細胞質送達とエンドソーム脱出促進。(左)200μg/mLのDex-488と10μME3MPH16を4℃もしくは37℃で3時間処理したHT1080細胞の共焦点顕微鏡画像。(右)左の画像において細胞質全体にDex-488シグナルが拡散した細胞の割合。E3MPH16による被送達物質の細胞内送達は4℃で抑制され、E3MPH16はエンドソーム経路から被送達物質を送達することが判明した。
E3MPH16の細胞質送達に関わる細胞内取り込み経路。(左)20nMコンカナマイシンで処置したのちに、20nMコンカナマイシン、10μME3MPH16と200μg/mLDex-488を添加し、3時間インキュベートしたHT1080細胞の共焦点顕微鏡画像。(右)左の画像において細胞質全体にDex-488シグナルが拡散した細胞の割合。E3MPH16による被送達物質の細胞内送達にはエンドソームの酸性化が重要であることが判明した。
E3MPH16の細胞質送達に関わる細胞内取り込み経路。各種Rab5変異体を発現させたHT1080細胞に対して10μME3MPH16と200μg/mLDex488を処理した際の共焦点顕微鏡画像。Rab5Q79Lを発現させても、Dex-488の細胞質送達に影響はほとんど見られなかったが、Rab5S34Nを発現させることでDex-488の細胞質送達がほとんど見られなくなった。E3MPH16による被送達物質の細胞内送達には初期エンドソームが重要であることが判明した。初期エンドソームへの移行を阻害すると、被送達物質が細胞質内で拡散できなくなることが判明した。
HT1080細胞におけるE3MPH16(Alexa)とリソソームマーカーLysoTracker redの共局在。10μMのE3MPH16(Alexa)を1時間インキュベートした後、HT1080細胞をLysotracker redで染色し、共焦点顕微鏡で生細胞画像を得た。E3MPH16(Alexa)はリソソームに局在化した。
MP(空白のダイヤモンド;◇)、E3MP(空白の三角形;△)、およびE3MPH16(塗りつぶしの円形;●)によって誘発されるリポソームリーケージアッセイ。CM(左)およびEM(右)は、細胞およびエンドソーム膜を模倣したリポソームを示す。E3MPH16は酸性脂質に対して障害性を発揮することが判明した。
異なる酸性アミノ酸をN末端に導入した時の影響。10μMの指定されたペプチドを200μg/mLのDex-488とともに18時間処理したHT1080細胞の共焦点顕微鏡画像。E3MPH16のN末端の3個のグルタミン酸をアスパラギン酸(D)または2-アミノアジピン酸(Aad)に置換しても、細胞内送達能は維持された。
H16MPE3のHT1080細胞における細胞毒性。H16MPE3を記載の濃度で18時間HT1080細胞に処理した際の細胞毒性。H16MPE3は100μMでも細胞毒性を示さなかった。
H16MPE3によるDex-488の細胞質送達。(上)200μg/mLDex-488と10μMの記載のペプチドで18時間処理したHT1080細胞の共焦点顕微鏡画像。(下)上の画像において細胞質でのDex-488シグナルを有する細胞の割合。H16MPE3はE3MPH16と同等に蛍光標識されたデキストランを細胞質に送達した。
【発明を実施するための形態】
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
国立大学法人京都大学
バイオセンサ
25日前
国立大学法人京都大学
多環芳香族化合物
17日前
国立大学法人京都大学
2次元フォトニック結晶レーザ
10日前
国立大学法人京都大学
2次元フォトニック結晶レーザ
19日前
国立大学法人京都大学
フルオロリン酸イオンを分離する方法
9日前
DMG森精機株式会社
表面発光装置
26日前
国立大学法人京都大学
BRG1遺伝子を標的とした膵癌治療剤
18日前
国立大学法人京都大学
車両管理システム、車両管理方法、及びプログラム
1日前
国立大学法人京都大学
NV中心作製方法およびダイヤモンド粒子の製造方法
16日前
国立大学法人京都大学
選別精子集団の製造方法及び男性不妊症の診断補助方法
1日前
DMG森精機株式会社
付加加工用ヘッドおよび加工機械
26日前
国立大学法人京都大学
コンピュータプログラム、真偽検証装置及び真偽検証方法
26日前
住友化学株式会社
高分子化合物、組成物、及び発光素子
24日前
三菱重工業株式会社
計画システム、計画方法およびプログラム
29日前
三菱重工業株式会社
計画システム、計画方法およびプログラム
29日前
住友金属鉱山株式会社
二酸化炭素還元光触媒粒子及びその製造方法
22日前
住友金属鉱山株式会社
二酸化炭素還元光触媒粒子及びその製造方法
16日前
住友金属鉱山株式会社
二酸化炭素還元光触媒粒子及びその製造方法
22日前
国立大学法人京都大学
バイオマス長尺体、複合長尺体、長尺状成形品およびバイオマス長尺体の製造方法
24日前
ジーエルサイエンス株式会社
ガラス瓶及びガラス瓶を用いたガス成分の分析方法
26日前
エスケーマテリアルズジェイエヌシー株式会社
多環芳香族化合物および有機電界発光素子
10日前
DMG森精機株式会社
付加加工用ヘッドおよび加工機械
26日前
国立大学法人京都大学
クラスター錯体およびその製造方法、そのクラスター錯体を構成ユニットとして有する高分子触媒およびその製造方法、ならびにその高分子触媒を含む電極およびその製造方法
17日前
東ソー株式会社
タンパク質の発現方法
8日前
東ソー株式会社
炭素-窒素結合形成方法
2か月前
株式会社トクヤマ
四塩化炭素の製造方法
2か月前
株式会社トクヤマ
シロキサン類の回収方法
2か月前
株式会社トクヤマ
ビオチン誘導体の製造方法
2か月前
株式会社半導体エネルギー研究所
有機化合物
1か月前
株式会社トクヤマ
ベンザゼピン化合物の製造方法
1か月前
株式会社コスモス
液状炭化水素の増産方法
1か月前
株式会社トクヤマ
ホルムアミド化合物の製造方法
1か月前
株式会社コスモス
液状炭化水素の増産方法
1か月前
東ソー株式会社
イソシアネート化合物の製造方法
1か月前
日本特殊陶業株式会社
メタン製造装置
3か月前
信越化学工業株式会社
新規化合物
2か月前
続きを見る
他の特許を見る

 特許ウォッチ
特許ウォッチ