TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025116476
公報種別
公開特許公報(A)
公開日
2025-08-08
出願番号
2024010920
出願日
2024-01-29
発明の名称
配座固定アミロイドβ42誘導体
出願人
国立大学法人京都大学
,
国立大学法人滋賀医科大学
代理人
個人
主分類
C07K
14/47 20060101AFI20250801BHJP(有機化学)
要約
【課題】ヒトの野生型アミロイドβ42と同じ毒性ターン構造を特異的に認識する抗体の開発や、超早期アルツハイマー病の迅速診断用試薬、毒性配座を有するアミロイドβ42に結合してその毒性を緩和する化合物、抗体医薬の開発に用いることができるアミロイドβ42誘導体を提供する。
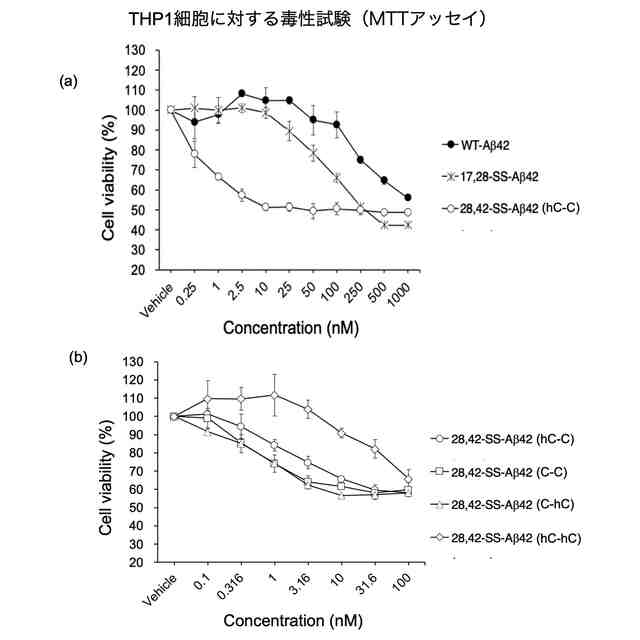
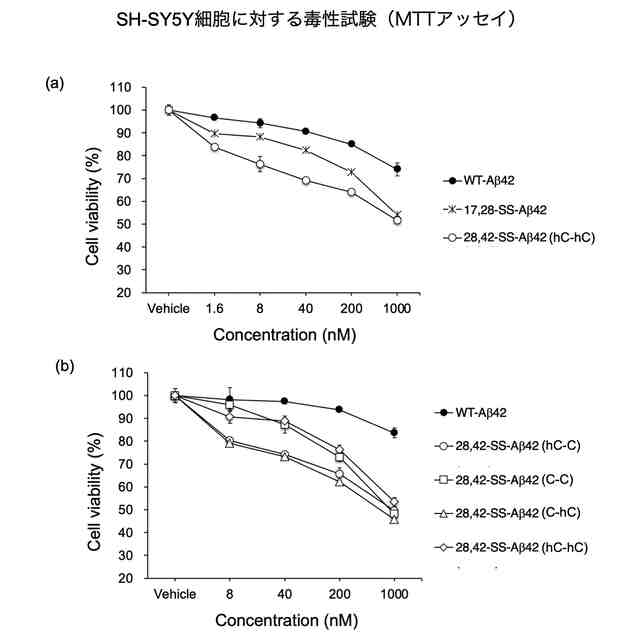
【解決手段】28位のアミノ酸残基と42位のアミノ酸残基が架橋され、22位のグルタミン酸残基と23位のアスパラギン酸残基においてアミノ酸配列によるβシート構造間のターン構造が固定された立体構造を有し、28位と42位以外のアミノ酸配列が、野生型アミロイドβ42と同じアミノ酸配列を有する配座固定アミロイドβ42誘導体を設計、合成したところ、極めて高い神経細胞毒性と、即時且つ持続的にβシート構造形成及び凝集能を有する誘導体が得られた。
【選択図】図4
特許請求の範囲
【請求項1】
アミノ酸42残基のアミロイドβ42において、
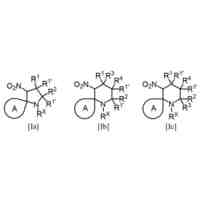
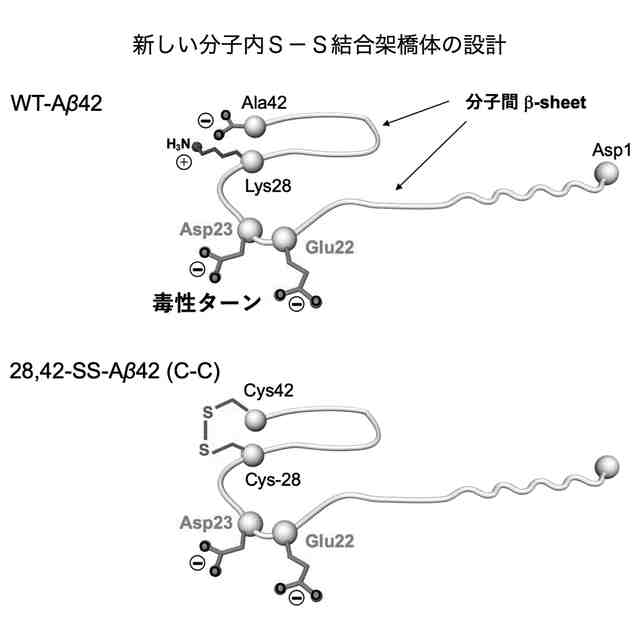
28位のアミノ酸残基と42位のアミノ酸残基が架橋され、βシート形成領域間に存在する22位のグルタミン酸残基と23位のアスパラギン酸残基からなるターン構造が固定された立体配座を有し、前記28位と前記42位のアミノ酸残基を除く全てのアミノ酸配列が、野生型アミロイドβ42と同じアミノ酸配列を有することを特徴とする配座固定アミロイドβ42誘導体。
続きを表示(約 490 文字)
【請求項2】
28位のリシン残基がシステイン残基又はホモシステイン残基に置換され、42位のアラニン残基がシステイン残基又はホモシステイン残基に置換され、28位と42位のアミノ酸残基間で分子内ジスルフィド結合を形成することにより、前記ターン構造が固定された請求項1に記載の配座固定アミロイドβ42誘導体。
【請求項3】
28位のリシン残基がホモシステイン残基に置換され、42位のアラニン残基がシステイン残基に置換された請求項2に記載の配座固定アミロイドβ42誘導体。
【請求項4】
28位のリシン残基がシステイン残基に置換され、42位のアラニン残基がホモシステイン残基に置換された請求項2に記載の配座固定アミロイドβ42誘導体。
【請求項5】
28位のリシン残基及び42位のアラニン残基が共にシステイン残基に置換された請求項2に記載の配座固定アミロイドβ42誘導体。
【請求項6】
28位のリシン残基及び42位のアラニン残基が共にホモシステイン残基に置換された請求項2に記載の配座固定アミロイドβ42誘導体。
発明の詳細な説明
【技術分野】
【0001】
本発明は、野生型のアミロイドβ42ペプチドの配列において毒性ターン構造を維持するように分子内で架橋し固定したアミロイドβ42の誘導体に関するものである。
続きを表示(約 5,400 文字)
【背景技術】
【0002】
アルツハイマー病(Alzheimer's disease:以下、「AD」という)は、神経変性疾患の一種であるが、現状では未だ有効な治療方法がほとんどなく、ADの発症機構の解明並びに根本的治療法の確立が強く望まれている。近年、ADの発症において、老人斑に蓄積するアミロイドβタンパク質(以下、「Aβ」という)が重要な役割を果たしており、その凝集により神経細胞毒性を示すことが判明してきている。ADでは、主に40残基または42残基のアミノ酸からなるAβ(以下、それぞれ、「Aβ40」と「Aβ42」という。)が蓄積することが特徴であり、ADの病因となることが見いだされているが、ADの発症においては、Aβ42がその凝集性や神経毒性の高さから、Aβ40よりも重要な役割を担っていると考えられている。しかしながら、抗Aβ抗体や抗タウ抗体等のADの治療薬はこれまでのところ成功例がほとんどなく、現在のところでは、進行を遅らせる医薬品(アリセプト、メマリー等)が承認されているのみである。このことについては、病気が進行してから治療を開始していることが失敗の原因である可能性があり、早期に診断を行い、適切な治療を開始できれば、ADを治療できる可能性があると考えられる。当初は、老人斑(Aβ凝集体)周辺の神経細胞が死滅していることなどから、Aβ凝集体を除去することがAD治療に有効と考えられていたが、老人斑(Aβ凝集体)の量とAD病態があまり相関しないことから、Aβ凝集体ではなく、準安定なAβオリゴマーが毒性本体であると考えられるようになってきた。ここで、「準安定」であるとは、質量分析等によりAβオリゴマーの構造を確認することができる時間、あるいは細胞を用いた試験においてAβオリゴマーの毒性を評価することができる時間として、例えば24時間程度はその状態で存在できることを意味している。ところが、数多くのAβオリゴマー種のうち、どれが細胞毒性に関わっているのか、またそれらがどのような高次構造をとっているのかについては、ほとんど明らかになっていない。さらに、これらのAβオリゴマーは、モノマーとアミロイド線維(フィブリル)との平衡状態にあり、純粋なオリゴマーを単離・構造決定することは不可能である。
【0003】
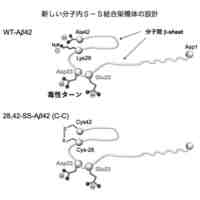
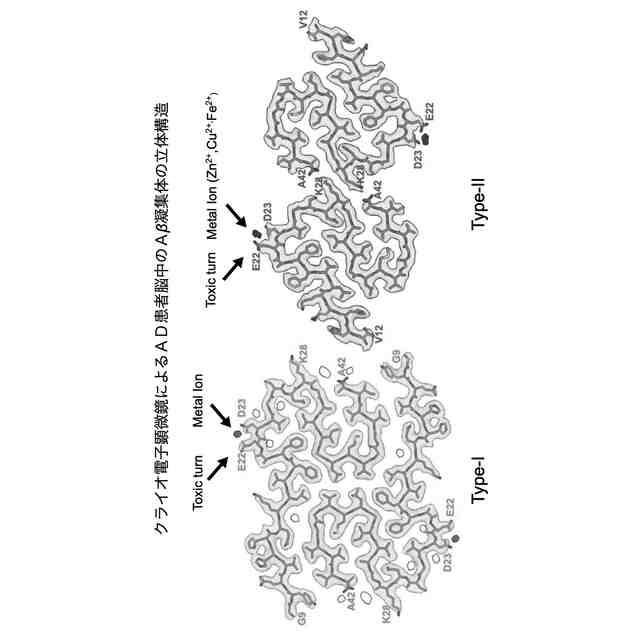
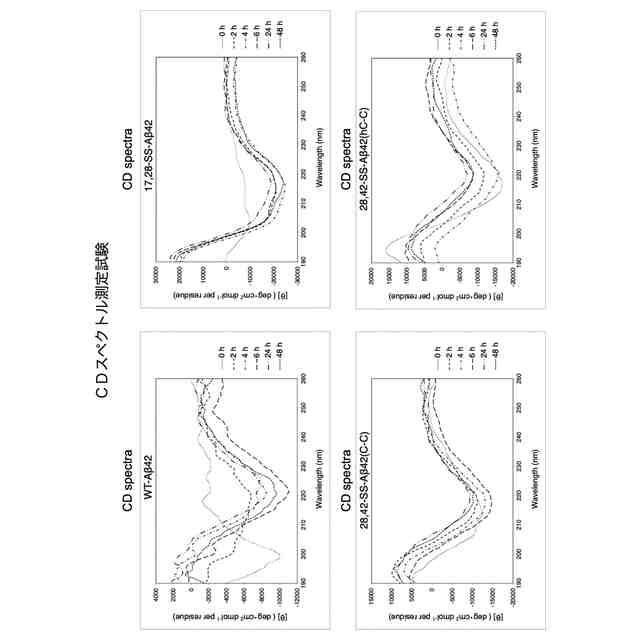
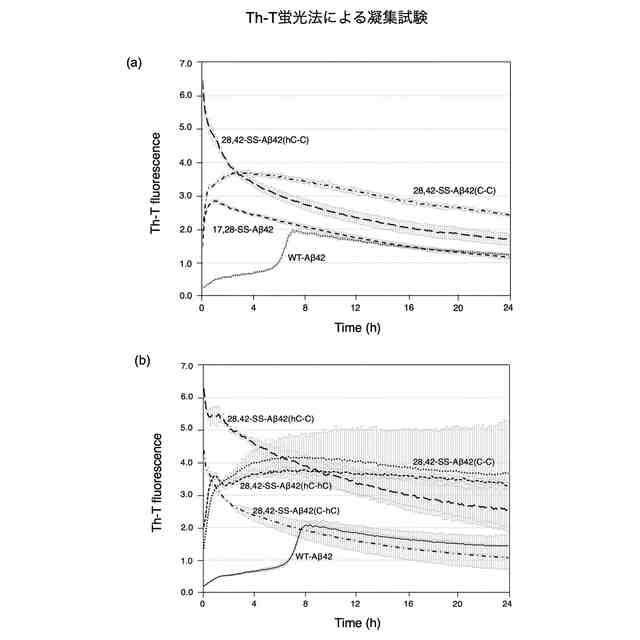
Aβのオリゴマー種の一部は、AD発症の初期段階でシナプス毒性を示すとともに、最終的に神経細胞の脱落をもたらすタウタンパク質の過剰リン酸化を誘発するために特に重要であると考えられている。本発明者の入江らはこれまでに、Aβ42の系統的なプロリン置換法、固体NMR法、電子スピン共鳴法などによる解析から、アミノ酸配列の中央部分[22位のグルタミン酸残基(E22)及び23位のアスパラギン酸残基(D23)の近傍]でのβシート構造間のターン構造(細胞毒性発現の原因となることから以下、「毒性ターン構造」とも称する。図1左上)とC末端の疎水性コア(図1右上)を特徴とした2量体及び3量体(図1下)のモデルを提唱している。すなわち、Aβのオリゴマー種のうち、毒性ターン構造をもつものや、一定以上の分子量(例えば、20量体以上)をもつものが、毒性発現に必要であると考えている(ACS Chem. Neurosci. 2017, 8 (4), 807-816)。さらに入江らは、特にAβ42の分子間βシート構造間の中央部分(22位及び23位の近傍)で折れ曲がることにより毒性オリゴマーを形成する(図1右下)という「毒性配座理論」(非特許文献1)を提唱している。ここで、「毒性」とは、細胞に対する増殖抑制活性があること、細胞が死滅すること、長期増強(LTP)の抑制(シナプス毒性又は神経毒性)があること等を意味する。ヒトの野生型Aβ42(WT-Aβ42)のアミノ酸配列は配列表の配列番号1及び図1に示した通りである。
【0004】
毒性配座理論では、Aβ42の中央部分でのターン構造(特に、22位及び23位のアミノ酸残基を中心とした20位から25位のアミノ酸配列により構成される)の形成は、銅イオン等との反応によって生じた10位のチロシン残基(Y10)のフェノキシラジカルによる35位のメチオニン残基(M35)の硫黄原子の酸化を効率的に促進し、生じたカチオンラジカルをC末端のカルボキシラートアニオンにより安定化できるものと理解できる。その結果、電荷を失ったAβ42のC末端コアの疎水性が増大して凝集しやすくなり、2量体及び3量体のオリゴマーが形成される。一方で、Aβ42のC末端の中性ラジカルはオリゴマー内に存在することから、長期間に亘って酸化ストレスを細胞に与えることが可能となるものと考えられる。Aβ42は25位のグリシン残基(G25)と26位のセリン残基(S26)でもターン構造を形成する(図1右上)が、22位(E22)と23位(D23)におけるターン構造を形成したAβ42よりも遙かに毒性が低いことが判明しているため、細胞毒性には22位と23位における毒性ターン構造が大きく関与していると考えられる。毒性ターン構造では、E22及びD23のカルボキシ基が共に主鎖に対して外側を向く(シス配置)を取っている。このような研究を通じて、入江らは、Aβ42の凝集と神経細胞毒性の発現において鍵となるAβ42の構造因子を明らかにすることによって、有害と考えられる立体配座に固定されたAβ42の誘導体を開発してきている(特許文献1参照)。
【0005】
入江らはこれまでに、Aβ42のE22をプロリン置換した9残基目から35残基目からなるAβペプチド(E22P-Aβ9-35)をキャリアータンパク質と結合した誘導体を作製し、それを抗原として立体構造特異抗体である24B3抗体(「IBL-102抗体」ともいう。)を開発している。24B3抗体は、開発時には、毒性ターン構造を有するAβ42を特異的に認識すると考えられた抗アミロイドβモノクローナル抗体である(特許文献2参照)。24B3抗体は、E22P-Aβ及びその2量体モデルには強く結合するが、毒性ターン構造をあまりとっていないWT-Aβ42モノマーに対する結合能が低いモノクローナル抗体であり、ヒト脳髄液を用いたAD診断において一定の成功を収めている(非特許文献1、非特許文献2参照)。24B3抗体と既存のN末端抗体とのサンドイッチELISA法を入江らと免疫生物学研究所とにより共同開発し、京都府立医科大学の徳田隆彦教授(現量子科学技術研究開発機構)の協力によりAD患者の脳脊髄液を解析した結果、AD患者において毒性コンホマー量(毒性ターンをもつAβ分子種の量)の全Aβ42量に対する割合が、ADでない人と比べて有意に高いことが判明している。また、特発性正常圧水頭症患者においてもこの割合が高い傾向にあることが判明するとともに、ADへの移行を判断する一つの指標になり得ることが示唆された(順天堂大学・中島円准教授との共同研究)。さらに24B3抗体は、Aβ42の神経細胞毒性を数種の市販抗体よりも強く抑制したことから、入江らは千葉大学医学研究科と共同で、ADモデルマウス(Tg2576)に対する治療効果を検討した結果、高架式十字迷路並びに巣作り試験において、認知機能改善効果が認められた。24B3抗体を用いたサンドイッチELISAは、2016年11月に、免疫生物学研究所より研究用試薬として販売され、国内外で多くの研究者に使用されている。
【0006】
その後のX線共結晶構造解析の結果、24B3抗体は、上述のとおり毒性ターン構造をもつと考えられたE22P-Aβ誘導体及びその2量体モデルを特異的に認識して結合することが分かったが、生体内におけるWT-Aβ42の毒性ターン構造に対する結合様式はまったく不明なままである。これは、ヒト脳内には存在しないプロリン置換体であるE22P-Aβ誘導体を抗原(ハプテン)として用いていることも一因である。このことから、AD診断、特に超早期の診断により有用な抗体を開発するためには、抗原として、毒性ターンの位置でのプロリン置換または22位及び23位側鎖間のラクタム形成を行うことなく、毒性ターンの位置でWT-Aβ42のアミノ酸配列をもつAβ42誘導体を開発することが望ましいと考えられた。
【0007】
それに続く研究において、入江らは、E22とD23を中心として2箇所をシステイン残基(C)で置換した誘導体を系統的に合成したのち,それぞれをDMSO酸化して分子内でジスルフィド(S-S)結合を形成させた。その結果、17位のロイシン残基(L17)と28位のリシン残基(K28)を共にシステイン残基に置換して分子内S-S結合を形成させた架橋構造を持つAβ42誘導体[L17C,K28C-17,28-S-S-Amyloid-β42、以下、「17,28-SS-Aβ42(C-C)」(配列番号2)と表記する]を開発した。以下、分子内S-S結合により毒性ターン構造を固定した架橋構造を有するAβ42誘導体を、「SS-Aβ42」と表記する。この17,28-SS-Aβ42(C-C)は生体内で毒性を示すと考えられるWT-Aβ42の毒性ターン構造を形成するE22及びD23とその前後各4残基(18位~27位)がWT-Aβ42のアミノ酸配列と同じであって、特にE22が生体内に存在しないプロリン置換されておらず、またE22及びD23の側鎖間にラクタムを構築することなく毒性ターンの立体構造が安定的に固定されたものである。17,28-SS-Aβ42(C-C)[17,28-SS-Aβ42]は、それまで高い凝集性を示し最も神経細胞毒性が高かったE22P-Aβ42と同等以上の凝集性と神経細胞毒性を示すものであることが分かった(特許文献3、非特許文献3参照)。また、17,28-SS-Aβ(C-C)以外の誘導体として、L17をホモシステイン残基に置換しK28をシステイン残基に置換して分子内S-S結合を形成させた誘導体[17,28-SS-Aβ(hC-C)]や、L17をシステイン残基に置換しK28をホモシステイン残基に置換して分子内S-S結合を形成させた誘導体[17,28-SS-Aβ(C-hC)]も、高い凝集性と神経細胞毒性を示すことが分かった(特許文献4参照)。以下、17位と28位との間で分子内S-S結合を形成したAβ42誘導体を総称する場合、「17,28-SS-Aβ42」と表記する。
【先行技術文献】
【特許文献】
【0008】
特開2006-265189号公報
WO2016/092865国際公開公報
特開2021-123551(特許第7357354号)
特開2022-155700
【非特許文献】
【0009】
Irie,K., Biosci.Biotechnol.Biochem.2020,84(1),1-16.
入江一浩、有機合成化学協会誌、2019,77(12),1201-1208.
Y.Matsushima,et.al., Chemical Communications,2020,56(29),4118-4121
【発明の概要】
【発明が解決しようとする課題】
【0010】
ところで、固体NMR法による凝集体の立体構造解析から、図2に示すように、WT-Aβ42では28位のリジン残基(K28)と42位のアラニン残基(A42)とが分子内イオン結合することによって、C末端側が22位と23位での毒性ターン構造とは反対側に折れ曲がった立体構造となっており、この構造で分子間βシート構造を形成している(Xiao, Y. et al., Nat. Struct. Mol. Biol. 2015, 22, 499-505. Colvin, M. T. et al., J. Am. Chem. Soc. 2016, 138, 9663-9674. Walti, M. A. et al., Proc. Natl. Acad. Sci. USA 2016, 113, E4976-E4984. なお、Waltiの”a”は変母音)。一方、同図に示すように、WT-Aβ40では、WT-Aβ42と比較してC末端側が2残基分短いことから、28位のリジン残基のアミノ基が40位のバリン残基のカルボキシ基と分子内イオン結合することがなく、代わりに23位のアスパラギン酸残基のカルボキシ基と分子内イオン結合を形成している。その結果、C末端側が直線的に伸びた構造となっており、この構造で分子間βシート構造を形成している(Petkova, A. T. et al., Proc. Natl. Acad. Sci. USA 2002, 99, 16742-16747.)。このような立体構造の違いから、WT-Aβ42に毒性ターン構造が存在することが強く支持され、ほとんど毒性を示さないWT-Aβ40が毒性ターン構造を取りにくい理由が明らかとなった。
(【0011】以降は省略されています)
特許ウォッチbot のツイートを見る
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
国立大学法人京都大学
抗がん剤
1か月前
国立大学法人京都大学
バイオセンサ
25日前
国立大学法人京都大学
マグネトロン
4か月前
国立大学法人京都大学
多環芳香族化合物
1か月前
国立大学法人京都大学
多環芳香族化合物
1か月前
国立大学法人京都大学
多環芳香族化合物
17日前
国立大学法人京都大学
細胞質送達ペプチド
5か月前
国立大学法人京都大学
凍結細胞の製造方法
3か月前
国立大学法人京都大学
高さ調整可能な展示台
3か月前
個人
肝疾患の検出方法
3か月前
国立大学法人京都大学
剪断流を発生させる器具
5か月前
国立大学法人京都大学
電気伝導体及びその設計方法
1か月前
国立大学法人京都大学
ブロック共重合体の製造方法
2か月前
国立大学法人京都大学
2次元フォトニック結晶レーザ
19日前
国立大学法人京都大学
2次元フォトニック結晶レーザ
10日前
国立大学法人京都大学
データ読み出し方法および装置
3か月前
国立大学法人京都大学
陽極およびアルカリ水電解装置
2か月前
国立大学法人京都大学
配座固定アミロイドβ42誘導体
1か月前
国立大学法人京都大学
神経変性疾患の予防または治療薬
2か月前
国立大学法人京都大学
全固体イオン選択性電極用組成物
3か月前
国立大学法人京都大学
二酸化炭素を回収する方法および装置
1か月前
DMG森精機株式会社
表面発光装置
26日前
国立大学法人京都大学
フーリエ変換装置および無線通信装置
3か月前
国立大学法人京都大学
フルオロリン酸イオンを分離する方法
9日前
国立大学法人京都大学
BRG1遺伝子を標的とした膵癌治療剤
18日前
株式会社構造計画研究所
領域変形装置
1か月前
日本電信電話株式会社
細胞培養デバイス
3か月前
多摩川精機株式会社
圧電型角速度センサ
4か月前
日本電信電話株式会社
細胞培養デバイス
3か月前
日本電信電話株式会社
細胞培養デバイス
3か月前
日本電信電話株式会社
細胞培養デバイス
3か月前
東ソー株式会社
がんモデル動物の作製方法
1か月前
国立大学法人京都大学
含窒素スピロ環化合物の製法及びその化合物
5か月前
スタンレー電気株式会社
面発光レーザ装置
1か月前
国立大学法人京都大学
β-グルコシル配糖体の製造方法及び形質転換体
3か月前
大和ハウス工業株式会社
身体機能評価システム
5か月前
続きを見る
他の特許を見る




 特許ウォッチ
特許ウォッチ