TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025127979
公報種別
公開特許公報(A)
公開日
2025-09-02
出願番号
2024025021
出願日
2024-02-21
発明の名称
スフィンゴミエリン製造方法
出願人
株式会社シャローム
代理人
個人
主分類
C12N
15/12 20060101AFI20250826BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】植物を用いてスフィンゴミエリンを安価に大量に製造する方法を提供する。
【解決手段】植物細胞で発現可能なプロモーター領域の下流に、下記(a)から(e)のいずれかに記載のヒト由来のDNAを機能的に連結させたベクター。
(a)特定のアミノ酸配列からなるタンパク質をコードするDNA
(b)特定の塩基配列のコード領域を含むDNA
(c)前記特定のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)前記特定のアミノ酸配列と90%以上の同一性を有し、前記特定のアミノ酸配列からなるタンパク質と同等の機能をするタンパク質をコードするDNA
(e)(a)、(c)または(d)のいずれか1のDNA配列のコドンを植物に最適化したDNA
【選択図】なし
特許請求の範囲
【請求項1】
植物細胞で発現可能なプロモーター領域の下流に、下記(a)から(e)のいずれかに記載のDNAを機能的に連結させたベクター。
(a)配列番号:2に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号2に記載のアミノ酸配列と90%以上の同一性を有し、配列番号2に記載のアミノ酸配列からなるタンパク質と同等の機能をするタンパク質をコードするDNA。
(e)(a)~(d)のいずれか1のDNA配列のコドンを植物に最適化したDNA。
続きを表示(約 510 文字)
【請求項2】
請求項1に記載のベクターが導入された形質転換植物細胞。
【請求項3】
請求項2に記載の形質転換植物細胞から再生された、スフィンゴミエリンを生産する能力を有する植物体。
【請求項4】
請求項3に記載の植物体の子孫またはクローンである、スフィンゴミエリンを生産する能力を有する植物体。
【請求項5】
請求項3または4に記載のスフィンゴミエリンを生産する能力を有する植物体の繁殖材料。
【請求項6】
スフィンゴミエリンを生産する能力を有する植物体の製造方法であって、
(i)請求項1に記載のベクターを植物細胞に導入する工程、および
(ii)工程(i)においてベクターが導入された形質転換植物細胞から植物体を再生する工程、
を含む方法。
【請求項7】
請求項2に記載の形質転換植物細胞、もしくは請求項3または4に記載の植物体を用いることを特徴とする、スフィンゴミエリンの製造方法。
【請求項8】
動物由来または合成したスフィンゴミエリン合成酵素遺伝子を導入し、発現させた植物細胞。
発明の詳細な説明
【技術分野】
【0001】
本発明は、スフィンゴミエリンの製造方法に関する。より詳しくはスフィンゴミエリンの植物による製造方法に関する。
続きを表示(約 3,200 文字)
【背景技術】
【0002】
従来、スフィンゴミエリンは、牛の脳やニワトリ表皮などから安価に抽出されていた(例えば、特許文献1)。しかしながら、牛の脳の場合は狂牛病の危険があることから、他の生産方法の開発が求められていたが、未だ遺伝子組み換え技術等を用いた他生物種による生物生産の報告もない。
そこで、より安く大量にスフィンゴミエリンを製造する方法の開発が求められていた。
【先行技術文献】
【特許文献】
【0003】
特開2008-179588号公報
【発明の概要】
【発明が解決しようとする課題】
【0004】
植物を用いてスフィンゴミエリンを安価に大量に製造する方法を提供する。
【課題を解決するための手段】
【0005】
(1)植物細胞で発現可能なプロモーター領域の下流に、下記(a)から(e)のいずれかに記載のDNAを機能的に連結させたベクター。
(a)配列番号:2に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号:1に記載の塩基配列のコード領域を含むDNA
(c)配列番号:2に記載のアミノ酸配列において1または複数のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列からなるタンパク質をコードするDNA
(d)配列番号2に記載のアミノ酸配列と90%以上の同一性を有し、配列番号2に記載のアミノ酸配列からなるタンパク質と同等の機能をするタンパク質をコードするDNA。
(e)(a)~(d)のいずれか1のDNA配列のコドンを植物に最適化したDNA。
ここで「機能的に連結」とは、ヒト由来DNAがプロモーターから転写され、翻訳されて目的のタンパク質が発現するように連結されていることをいう。
(2)(1)に記載のベクターが導入された形質転換植物細胞。
(3)(2)に記載の形質転換植物細胞から再生された、スフィンゴミエリンを生産する能力を有する植物体。
(4)(3)に記載の植物体の子孫またはクローンである、スフィンゴミエリンを生産する能力を有する植物体。
(5)(3)または(4)に記載のスフィンゴミエリンを生産する能力を有する植物体の繁殖材料。
(6)スフィンゴミエリンを生産する能力を有する植物体の製造方法であって、
(i)(1)に記載のベクターを植物細胞に導入する工程、および
(ii)工程(i)においてベクターが導入された形質転換植物細胞から植物体を再生する工程、
を含む方法。
(7)(2)に記載の形質転換植物細胞、(3)または(4)に記載の植物体を用いることを特徴とする、スフィンゴミエリンの製造方法。
(8) 動物由来または合成したスフィンゴミエリン合成酵素遺伝子を導入し、発現させた植物細胞。ここで、合成とは、一部合成および全部合成も含む意味である。
【発明の効果】
【0006】
本発明によれば、動物由来の感染症のおそれがない安全な植物で、安価に大量に合成する方法が提供される。
【図面の簡単な説明】
【0007】
図1は、hSMS2遺伝子のDNA配列を示す図である(実施例1)。
図2は、hSMSタンパク質のアミノ酸配列を示す図である。
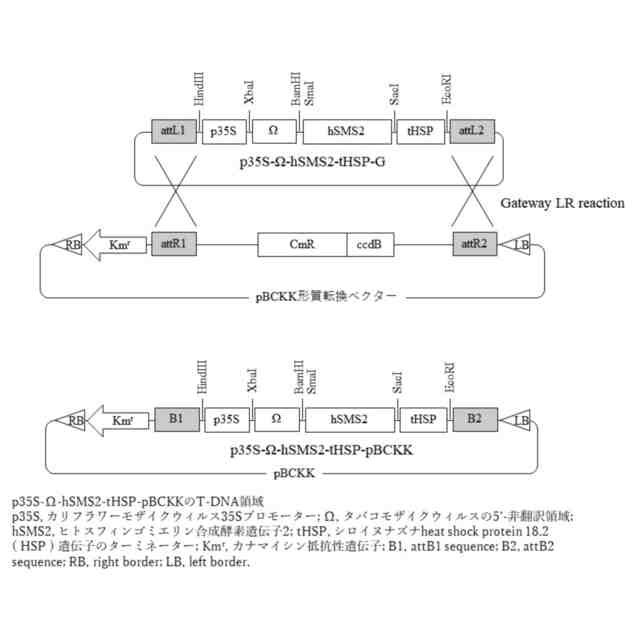
図3は、p35S-Ω-hSMS2-tHSP-pBCKKベクターの図である(実施例2)。
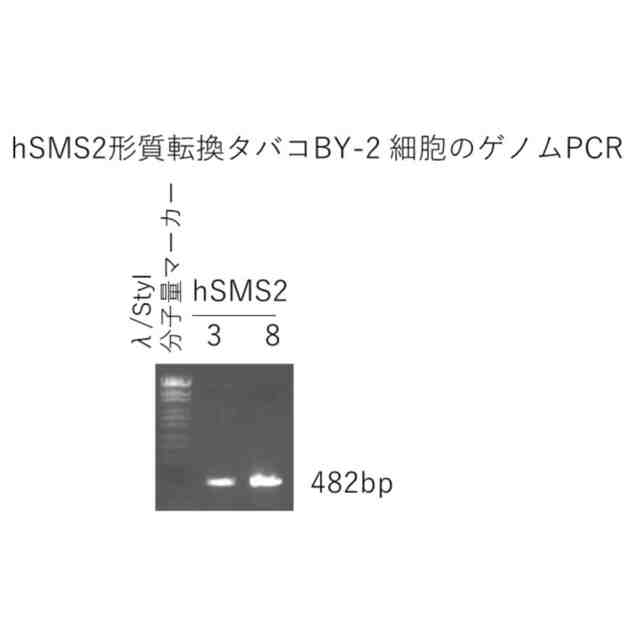
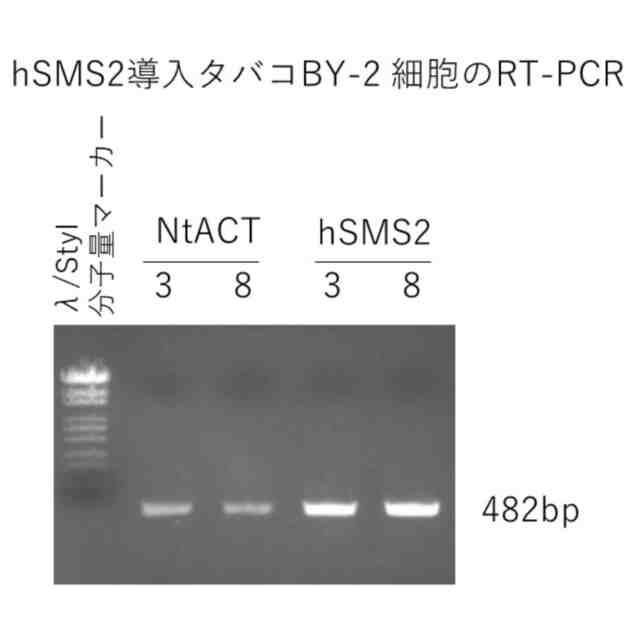
図4は、hSMS2形質転換タバコBY-2細胞のゲノムPCRの結果を示す写真である(実施例4)。
図5は、hSMS2形質転換タバコBY-2細胞のRT-PCRの結果を示す写真である(実施例5)。
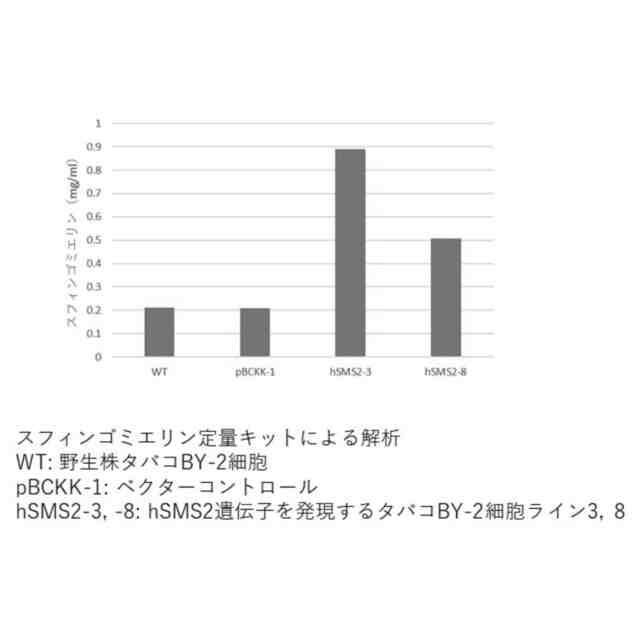
図6は、スフィンゴミエリン定量キットによる定量結果を示すグラフである(実施例7)。
図7は、LC-MS/MSによる網羅的脂質解析データを示す表と網羅的脂質解析データを用いてセラミド、グルコシルセラミド、スフィンゴミエリンを定量したグラフである(実施例8)。
図8は、hSMS1遺伝子のDNA配列を示す図である(実施例9)。
図9は、hSMS1 Art遺伝子のDNA配列を示す図である(実施例9)。
図10は、hSMS2 Art遺伝子のDNA配列を示す図である(実施例9)。
図11は、hSMS1 Art遺伝子を含むイネ用形質転換ベクターを示す図である(実施例13)。
図12は、イネEF1α-1プロモーター配列を示す図である(実施例12)。
図13は、EF1α-1またはEF1α-4プロモーター-hSMS2を含む形質転換ベクターの構造を示す図である(実施例12)。
図14は、イネ種子特異的Globulinプロモーター配列を示す図である(実施例13)。
図15は、Globulinプロモーター-hSMS1 Art-DT2を含む発現ベクターの構造を示す図である(実施例13)。
図16は、形質転換イネのゲノムPCRの結果を示す写真である(実施例14)。
図17は、形質転換イネの穀粒のRT-PCRの結果を示す写真である(実施例14)。
図18は、イネ形質転換体の網羅的脂質解析の結果を示すグラフである(実施例15)。
図19は、コドン最適化したSMS2遺伝子に35Sプロモーターを連結した形質転換ベクターの構成を示す図である(実施例15)。
図20は、コドン最適化hSMS2を導入したタバコBY-2細胞の発現量(A)とスフィンゴミエリン量(B)を測定したグラフである(実施例10)。
図21は、ELF1―α4プロモーターの塩基配列を示す図である(実施例12)。
図22は、コドン最適化したSMS2遺伝子を導入したイネ形質転換体のRT-PCRを示す写真である(実施例14)。
図23は、イネ形質転換体のスフィンゴミエリンの定量結果を示す図である(実施例16)。
【発明を実施するための形態】
【0008】
本発明は、スフィンゴミエリン合成酵素(SMS)遺伝子を植物細胞に導入、発現させることにより、植物細胞中でスフィンゴミエリンを生産することに特徴がある。SMSとしては、もともと植物にはスフィンゴミエリンが存在しないことから、外来遺伝子導入によりスフィンゴミエリンが製造できるかは不明であった。それに加えて、大腸菌など他生物種でのスフィンゴミエリン生産の報告例もない。
【0009】
本発明において、SMS遺伝子としては、ヒト由来SMS2遺伝子が好適に用いられるが、SMS1遺伝子であってもよい。SMS2、SMS1のDNA配列、アミノ酸配列は、図1、2,8に示す。また、配列番号1~4に示す(配列表参照)。また、SMS遺伝子として、SMS2タンパク質と90%以上の同一性を有し、SMS2タンパク質と同等のSMS機能を有するタンパク質をコードするDNAも用いることができる。SMS遺伝子の由来としては、ヒトに限らず、哺乳類、鳥類、は虫類、両生類、魚類、甲殻類、昆虫類などのSMS遺伝子を用いてもよい。
【0010】
また、hSMS2タンパク質のアミノ酸配列から1から数個(または複数個)アミノ酸が置換したアミノ酸配列からなるタンパク質であって、hSMS2タンパク質と同等の機能を有するタンパク質をコードするDNAも用いることができる。本明細書において、「同等の機能を有する」とは同程度の酵素活性を示すことをいう。
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
個人
抗遺伝子劣化装置
1か月前
個人
細胞内探査とその利用
1か月前
雪国アグリ株式会社
単糖類の製造方法
10日前
個人
細胞培養容器
3か月前
日本バイリーン株式会社
細胞用支持基材
3か月前
株式会社東洋新薬
経口組成物
1か月前
株式会社タクマ
バイオマス処理装置
2か月前
株式会社タクマ
バイオマス処理装置
2か月前
テルモ株式会社
吐出デバイス
1か月前
東洋紡株式会社
改変型RNAポリメラーゼ
3か月前
日油株式会社
蛋白質安定化剤
3か月前
島根県
油吸着材とその製造方法
8日前
宝酒造株式会社
アルコール飲料
19日前
テルモ株式会社
容器蓋デバイス
1か月前
東ソー株式会社
pH応答性マイクロキャリア
1か月前
大陽日酸株式会社
培養装置
1か月前
大陽日酸株式会社
培養装置
1か月前
トヨタ自動車株式会社
バイオ燃料製造方法
1か月前
株式会社ファンケル
SEC12タンパク発現促進剤
3か月前
株式会社東海ヒット
灌流培養ユニット
4か月前
個人
有機フッ素化合物を分解する廃液処理法
4日前
株式会社豊田中央研究所
細胞励起装置
17日前
アサヒビール株式会社
柑橘風味アルコール飲料
3か月前
アサヒビール株式会社
柑橘風味アルコール飲料
3か月前
オンキヨー株式会社
浸漬酒の製造方法、及び、浸漬酒
3か月前
テルモ株式会社
採取組織細切補助デバイス
1か月前
株式会社シャローム
スフィンゴミエリン製造方法
1か月前
株式会社村田製作所
濾過装置および濾過方法
9日前
株式会社今宮
瓶詰ビールの加熱殺菌方法および装置
1か月前
新東工業株式会社
培養システム
9日前
横河電機株式会社
藻類培養装置
29日前
花王株式会社
リパーゼ変異体
3か月前
住友金属鉱山株式会社
連続発酵方法及び連続発酵装置
1か月前
東ソー株式会社
免疫学的測定法
4日前
株式会社ショウワ
キトサンオリゴマー分画方法
19日前
公立大学法人北九州市立大学
微生物の検知方法
2か月前
続きを見る
他の特許を見る






 特許ウォッチ
特許ウォッチ