TOP
|
特許
|
意匠
|
商標
特許ウォッチ
Twitter
他の特許を見る
10個以上の画像は省略されています。
公開番号
2025143470
公報種別
公開特許公報(A)
公開日
2025-10-01
出願番号
2025116324,2021552482
出願日
2025-07-10,2020-10-16
発明の名称
グルクロン酸転移酵素、それをコードする遺伝子及びその利用方法
出願人
国立大学法人大阪大学
,
国立研究開発法人農業・食品産業技術総合研究機構
代理人
弁理士法人平木国際特許事務所
主分類
C12N
1/19 20060101AFI20250924BHJP(生化学;ビール;酒精;ぶどう酒;酢;微生物学;酵素学;突然変異または遺伝子工学)
要約
【課題】オレアナン型トリテルペノイドの3位のヒドロキシル基にグルクロン酸転移を触媒するグルクロン酸第1転移酵素の遺伝子を同定する。
【解決手段】マメ科植物(ダイズ、カンゾウ、及びミヤコグサ)に由来し、それぞれ配列番号2、4、及び6で示す塩基配列からなる目的の活性を有したグルクロン酸第1転移酵素遺伝子を提供する。
【選択図】なし
特許請求の範囲
【請求項1】
β-アミリンを生合成でき、かつ以下の(1)~(3)で示すポリペプチドをコードするポリヌクレオチドが全て導入された遺伝子組換え体。
(1)オレアナン型トリテルペノイドにおける11位を酸化する活性を有し、以下の(a
)又は(b)で示すいずれかのアミノ酸配列を含むポリペプチド、
(a)配列番号7で示すアミノ酸配列、又は
(b)配列番号7で示すアミノ酸配列と90%以上の同一性を有するアミノ酸配列、
(2)オレアナン型トリテルペノイドにおける30位を酸化する活性を有し、以下の(c
)又は(d)で示すいずれかのアミノ酸配列を含むポリペプチド、
(c)配列番号13で示すアミノ酸配列、又は
(d)配列番号13で示すアミノ酸配列と90%以上の同一性を有するアミノ酸配列、
(3)オレアナン型トリテルペノイドにおける3位のヒドロキシ基にグルクロン酸を転
移する活性を有し、以下の(e)又は(f)で示すいずれかのアミノ酸配列を含むポリペ
プチド、
(e)配列番号1、3、及び5のいずれかで示すアミノ酸配列、又は
(f)配列番号1、3、及び5のいずれかで示すアミノ酸配列と90%以上の同一性を
有するアミノ酸配列。
続きを表示(約 330 文字)
【請求項2】
さらに、以下の(4)で示すポリペプチドをコードするポリヌクレオチドが導入された、請求項1に記載の遺伝子組換え体。
(4)UDP-グルコースを基質としてUDP-グルクロン酸を生合成する活性を有するポリペプチド。
【請求項3】
前記(4)が、配列番号25で示す塩基配列によりコードされるポリペプチドである、請求項2に記載の遺伝子組換え体。
【請求項4】
宿主が酵母である、請求項1~3のいずれか一項に記載の遺伝子組換え体。
【請求項5】
β-アミリンからグリチルレチン酸モノグルクロニドを生合成する方法であって、請求項1~4のいずれか一項に記載の遺伝子組換え体を培養する工程を含む前記方法。
発明の詳細な説明
【技術分野】
【0001】
本発明は、オレアナン型トリテルペノイドにおける3位のヒドロキシル基にグルクロン酸を転移する酵素、当該酵素をコードする遺伝子、及びグリチルリチンの製造方法に関する。
続きを表示(約 4,800 文字)
【背景技術】
【0002】
カンゾウ(Glycyrrhiza uralensis:グリキルリザ ウラレンシス)は、マメ科の多年生草本植物である。この植物の根部及びストロン(stolon:地下茎)は、漢方上重要な生薬「甘草」として知られており、世界的に広く利用されている。甘草の主活性成分は、オレアナン型トリテルペノイドサポニンのグリチルリチン(glycyrrhizin)である(非特許文献1)。グリチルリチンについては、その生薬学的、薬理学的有用性、及び育種学的研究等の様々な側面から研究が行われている。
【0003】
医薬品として良質のグリチルリチンを生物生産系によって安定的かつ持続的に提供するためには、グリチルリチンの生合成系に関与する遺伝子や当該遺伝子の発現量をマーカーに用いて、最適産生条件の確立、グリチルリチン高生産株の選抜又は合成酵素遺伝子の導入によるグリチルリチン高産生植物の育種等を行う必要がある。そのためにはグリチルリチンの生合成系に関与する遺伝子群の同定が不可欠である。
【0004】
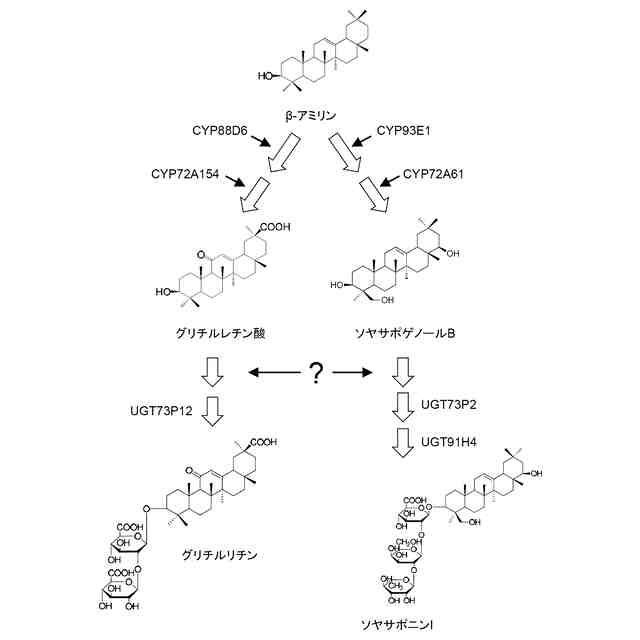
グリチルリチンは、植物に共通に含まれ、オレアナン型トリテルペノイドに属するβ-アミリンを出発物質として、2段階の酸化反応及び2段階の配糖化反応によって生合成される。β-アミリンとは、トリテルペノイドサポニン生合成系において、グリチルリチンとソヤサポニンI(soyasaponin I)の生合成分岐点となる前駆体物質であることが知られている(図1)。
【0005】
図2で示すように、β-アミリンからグリチルリチンに至る合成酵素は、これまでに、グリチルリチンのアグリコン(非糖部)であるグリチルレチン酸をβ-アミリンから生合成するための前記2段階の酸化反応のそれぞれを触媒する2種の酸化酵素CYP88D6(特許文献1)及びCYP72A154(特許文献2)、並びに得られたグリチルレチン酸に対して前記2段階の配糖化反応を触媒してグリチルリチンを生合成する合成酵素遺伝子のうち、2段階目を触媒する糖転移酵素UGT73P12(特許文献3)については知られていた。ところが、グリチルレチン酸に直接グルクロン酸を転移する1段階目の配糖化反応を触媒する糖転移酵素、すなわちグルクロン酸第1転移酵素については、多くの当業者が幾度となくその単離を試みたにもかかわらず、これまで全く得ることができなかった。それ故に、β-アミリンから生物生産系やインビトロ合成系でグリチルリチンを合成する上でのボトルネックとなっており、十分量のグリチルリチンを安定的かつ持続的に得ることができなかった。
【先行技術文献】
【特許文献】
【0006】
特許第5526323号
特許第5771846号
特許第6344774号
【非特許文献】
【0007】
Gibson, M. R., 1978, Lloydia-the journal of Natural Products, 41(4): 348-354
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、上記課題を解決するためにグリチルレチン酸をはじめとするオレアナン型トリテルペノイドの3位のヒドロキシル基にグルクロン酸転移を触媒するグルクロン酸第1転移酵素の遺伝子を単離すると共に、その遺伝子発現系を用いて個体内又は細胞内においてβ-アミリンからグリチルリチンまでを多量に生合成できる生物発現系を作製し、提供することを目的とする。
【課題を解決するための手段】
【0009】
β-アミリンからグリチルリチンへの生合成経路において、最後に残されたグルクロン酸第1転移酵素を同定するため、多くの当業者がカンゾウからの単離を試みたものの、長きにわたって同定されることができなかった。そこで、本発明者らは、カンゾウ属植物からのグルクロン酸第1転移酵素の単離には、何らかの阻害要因があると考え、カンゾウ属植物以外の植物からグルクロン酸第1転移酵素を単離する戦略を試みた。カンゾウ属植物が属するマメ科植物には、一般に、β-アミリンから中間産物としてソヤサポゲノールBを経由してソヤサポニンIを生合成する経路が存在する(図1)。オレアナン型トリテルペノイドであるソヤサポゲノールBは3位にヒドロキシル基を有するが、最終産物のソヤサポニンIは3位にはグルクロン酸が結合している。これは、中間産物でオレアナン型トリテルペノイドでもあるグリチルレチン酸が3位にヒドロキシル基を有するのに対して、最終産物のグリチルリチンの3位にはグルクロン酸が結合しているのと同じである。つまり、β-アミリンからグリチルリチンへの生合成経路で機能するグルクロン酸第1転移酵素は、β-アミリンからソヤサポニンIへの生合成経路でも機能している可能性がある。本発明者らは、この仮説に基づいて、β-アミリンからソヤサポニンIへの生合成経路を有するダイズからグルクロン酸第1転移活性を有し得る遺伝子の同定を試みた結果、具体的な機能が未知のセルロース合成酵素類似遺伝子を単離することに成功した。ソヤサポゲノールBを糖受容基質として、この酵素の糖転移活性を検証した結果、ソヤサポゲノールBの3位のヒドロキシル基にグルクロン酸を糖転移することが明らかとなった。この酵素活性は、糖受容基質がグリチルレチン酸の場合でも同様であった。このように、本発明者らは、由来植物をカンゾウ属植物ではなく、ダイズに変えたことで、これまで単離できなかったグルクロン酸第1転移酵素を同定することに成功した。本発明は、その研究結果に基づくものであり、以下を提供する。
【0010】
(1)オレアナン型トリテルペノイドにおける3位のヒドロキシ基にグルクロン酸を転移する活性を有し、以下の(a)~(c)で示すいずれかのアミノ酸配列を含むポリペプチド又は前記活性を有するその断片。
(a)配列番号1、3、及び5のいずれかで示すアミノ酸配列、
(b)配列番号1、3、及び5のいずれかで示すアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列、又は
(c)配列番号1、3、及び5のいずれかで示すアミノ酸配列と80%以上の同一性を有するアミノ酸配列
ポリペプチド。
(2)前記オレアナン型トリテルペノイドが、β-アミリン、11-オキソ-β-アミリン、30-ヒドロキシ-11-オキソ-β-アミリン、30-ヒドロキシ-β-アミリン、24-ヒドロキシ-β-アミリン、11-デオキソグリチルレチン酸、グリチルレチン酸、オレアノール酸、メジカゲニン酸、ソヤサポゲノールB、ソヤサポゲノールA、ヘデラゲニン、カメリアゲニン、及びサイコゲニンからなる群から選択される、(1)に記載のポリペプチド。
(3)マメ科(Fabaceae)植物に由来する、(1)又は(2)に記載のポリペプチド。
(4)(1)~(3)のいずれかに記載のポリペプチドをコードするポリヌクレオチド。
(5)以下の(a)~(d)で示すいずれかの塩基配列を含む、(4)に記載のポリヌクレオチド。
(a)配列番号2、4、及び6のいずれかで示す塩基配列、
(b)配列番号2、4、及び6のいずれかで示す塩基配列において1若しくは複数個の塩基が欠失、置換若しくは付加された塩基配列、
(c)配列番号2、4、及び6のいずれかで示す塩基配列と80%以上の同一性を有する塩基配列、又は
(d)配列番号2、4、及び6のいずれかで示す塩基配列に相補的な塩基配列と高ストリンジェントな条件でハイブリダイズする塩基配列
(6)(4)又は(5)に記載のポリヌクレオチドを含むCSyGT発現ベクター。
(7)(4)又は(5)に記載のポリヌクレオチド又は(6)に記載のCSyGT発現ベクターを含む形質転換体、又は前記ポリヌクレオチド又は前記CSyGT発現ベクターを保持したその後代。
(8)宿主がマメ科(Fabaceae)植物である、(7)に記載の形質転換体又はその後代。
(9)宿主が酵母である、(7)に記載の形質転換体又はその後代。
(10)(9)に記載の形質転換体又はその後代から得られる酵母由来の糖鎖が付加された(1)~(3)のいずれかに記載のポリペプチド。
(11)前記酵母由来の糖鎖が高マンノース型糖鎖である、(10)に記載のポリペプチド。
(12)オレアナン型トリテルペノイドにおけるグルクロン酸の2位のヒドロキシ基にグルクロン酸を転移する活性を有するポリペプチドを製造する方法であって、(7)又は(8)に記載の形質転換体又はその後代を培養する工程、及び前記培養物から(1)~(3)のいずれかに記載のポリペプチドを抽出する工程を含む、前記方法。
(13)β-アミリンを生合成でき、かつ以下の(A)~(D)で示す全ての発現ベクターを含むグリチルリチン製造用の遺伝子組換え体。
(A)オレアナン型トリテルペノイドにおける11位を酸化する活性を有し、以下の(a)~(c)で示すいずれかのアミノ酸配列を含むポリペプチドを包含するCYP88D6発現ベクター、
(a)配列番号7で示すアミノ酸配列、
(b)配列番号7で示すアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列、又は
(c)配列番号7で示すアミノ酸配列と80%以上の同一性を有するアミノ酸配列、
(B)オレアナン型トリテルペノイドにおける30位を酸化する活性を有し、以下の(d)~(f)で示すいずれかのアミノ酸配列を含むポリペプチドを包含するCYP72A154発現ベクター、
(d)配列番号9、11、及び13のいずれかで示すアミノ酸配列、
(e)配列番号9、11、及び13のいずれかで示すアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列、又は
(f)配列番号9、11、及び13のいずれかで示すアミノ酸配列と80%以上の同一性を有するアミノ酸配列、
(C)オレアナン型トリテルペノイドモノグルクロニドにおけるグルクロン酸の2位のヒドロキシ基にグルクロン酸を転移する活性を有し、以下の(g)~(i)で示すいずれかのアミノ酸配列を含むポリペプチドを包含するUGT73P12発現ベクター、
(g)配列番号15で示すアミノ酸配列、
(h)配列番号15で示すアミノ酸配列において1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列、又は
(i)配列番号15で示すアミノ酸配列と80%以上の同一性を有するアミノ酸配列、及び
(D)(6)に記載のCSyGT発現ベクター
(14)宿主がマメ科植物である、(13)に記載の遺伝子組換え体。
(15)β-アミリンからグリチルリチンを製造する方法であって、(13)又は(14)に記載の遺伝子組換え体を培養する工程を含む前記製造方法。
本明細書は本願の優先権の基礎となる日本国特許出願番号2019-190060号の開示内容を包含する。
【発明の効果】
(【0011】以降は省略されています)
この特許をJ-PlatPat(特許庁公式サイト)で参照する
関連特許
国立大学法人大阪大学
細胞シート
3か月前
国立大学法人大阪大学
モータ装置
7日前
国立大学法人大阪大学
内視鏡システム
4か月前
国立大学法人大阪大学
眼瞼痙攣緩和具
2か月前
国立大学法人大阪大学
半月板の治療薬
29日前
国立大学法人大阪大学
ロボットハンド
1か月前
国立大学法人大阪大学
漏洩情報抑制回路
2か月前
国立大学法人大阪大学
試料中の標的物質の検出
1か月前
国立大学法人大阪大学
化学反応方法及び反応装置
22日前
トヨタ自動車株式会社
電池
3か月前
国立大学法人大阪大学
電気泳動装置および電気泳動法
1か月前
大和ハウス工業株式会社
反応装置
10日前
大和ハウス工業株式会社
反応装置
10日前
川崎重工業株式会社
ゼオライト触媒
16日前
国立大学法人大阪大学
化合物の製造方法、及び金属多核錯体
1か月前
国立大学法人大阪大学
タンパク質間相互作用を可視化する方法
1か月前
国立大学法人大阪大学
溶融試料浮遊装置および溶融試料浮遊方法
8日前
国立大学法人大阪大学
ナノダイヤモンドの製造方法および製造装置
3か月前
株式会社トクヤマ
ケトン誘導体の製造方法
2か月前
セイコーグループ株式会社
生体電位計測装置
28日前
セイコーグループ株式会社
生体電位計測装置
28日前
株式会社アイシン
フッ化物イオン電池用電解液
7日前
株式会社塚田メディカル・リサーチ
カテーテル
4か月前
日東紡績株式会社
二酸化炭素の吸収又は脱着剤
2か月前
国立大学法人大阪大学
セミクラスレートハイドレートおよびその製造方法
4か月前
国立大学法人大阪大学
トラッキング装置、トラッキング方法及びプログラム
3か月前
ウシオ電機株式会社
半導体レーザ素子および発光装置
18日前
三菱重工業株式会社
劣化診断方法および劣化診断装置
4か月前
エヌ・イーケムキャット株式会社
還元的アミノ化触媒
2か月前
ウシオ電機株式会社
半導体レーザ素子および発光装置
14日前
日本電気株式会社
推定装置、推定方法及びプログラム
8日前
日東紡績株式会社
電池用活物質、電池用電極及び電池
9日前
NTT株式会社
量子鍵配送システム及び量子鍵配送方法
9日前
国立大学法人大阪大学
コンピュータプログラム、情報処理方法及び情報処理装置
9日前
株式会社塚田メディカル・リサーチ
バルーンカテーテル
4か月前
大成建設株式会社
汚泥処理方法および汚泥処理システム
1か月前
続きを見る
他の特許を見る









 特許ウォッチ
特許ウォッチ